森羅万象学校 2001
光合成生物への進化
山本 啓之, 川上 伸一
2001 年 9 月 12 日

|
表紙
|
|
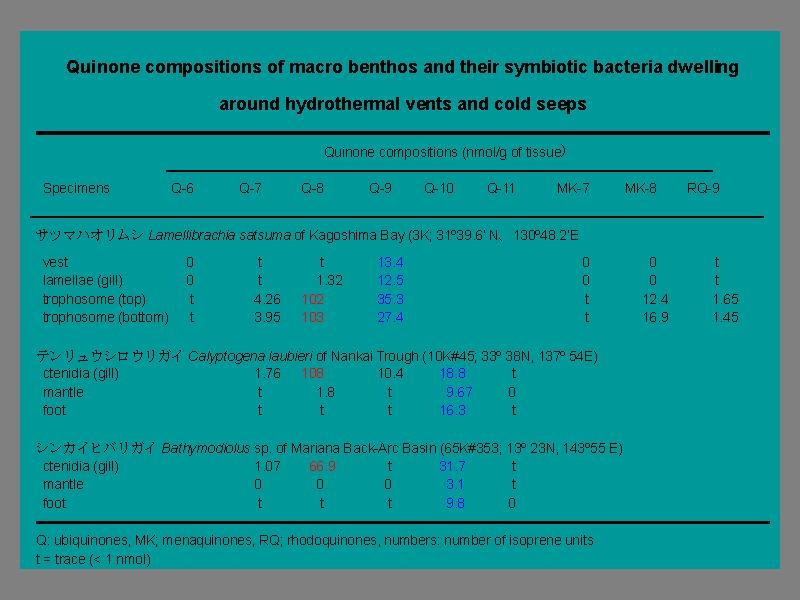
複数の貝類に対する染色物質の分布
- H2S かメタンハイドレードが利用できる環境で生息する貝について.
|

|
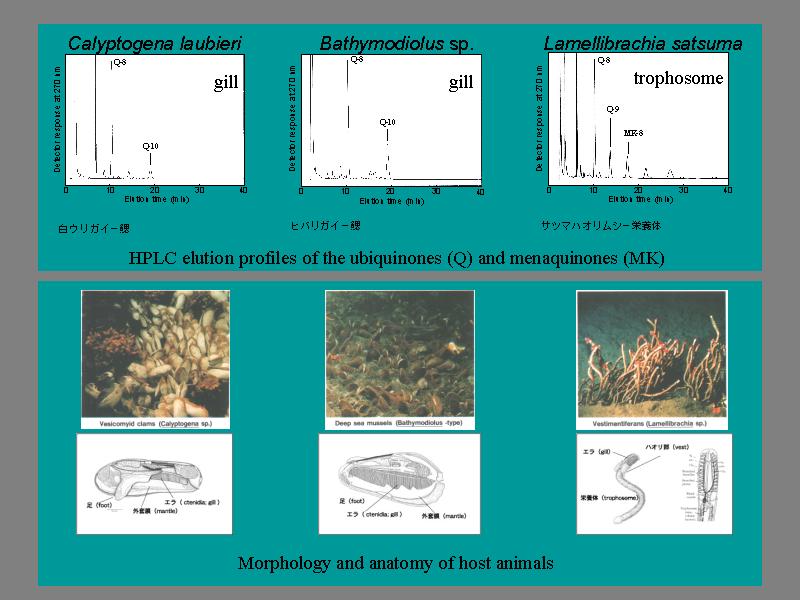
液体クロマトグラフィーの結果
共生細菌によるピークが現れる.
- シロウリガイ, ヒバリガイからはエラからのみ検出.
- ハオリムシは全身から検出される. 消化機能をバクテリアに完全に異存.
|

|
貝に共生するバクテリアの系統樹
- シロウリガイは親か子へ渡す
- ヒバリガイ, ハオリムシは周囲の環境から取り込む
|
|
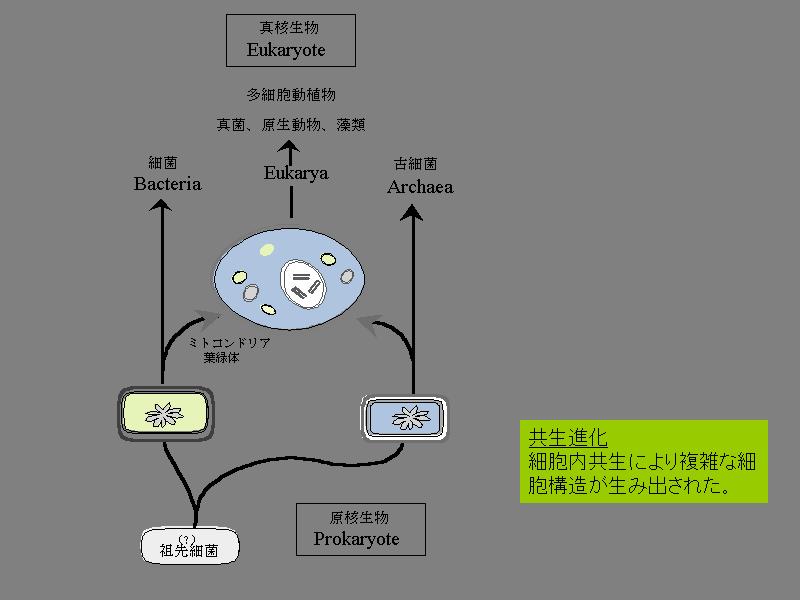
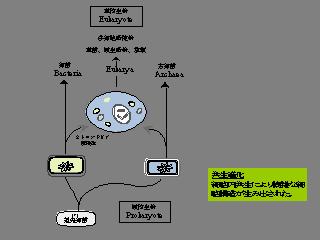
共生進化の模式図
|

|
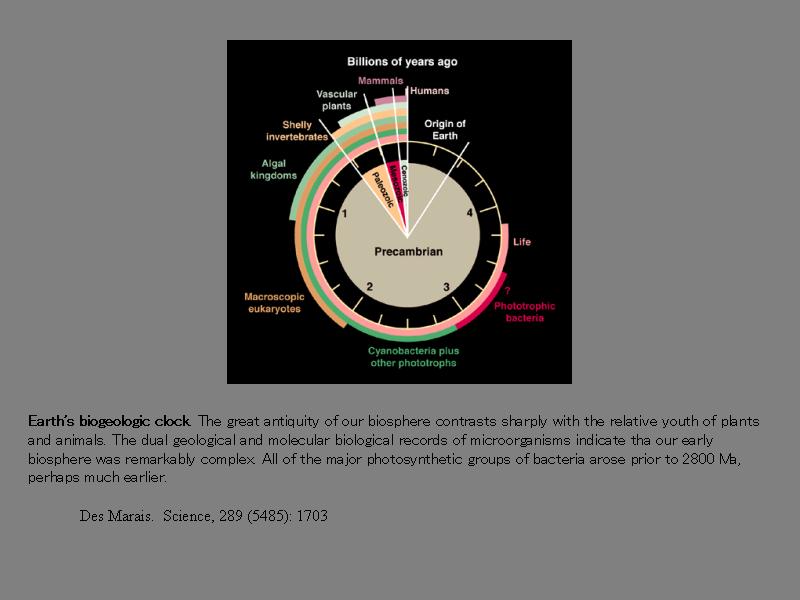
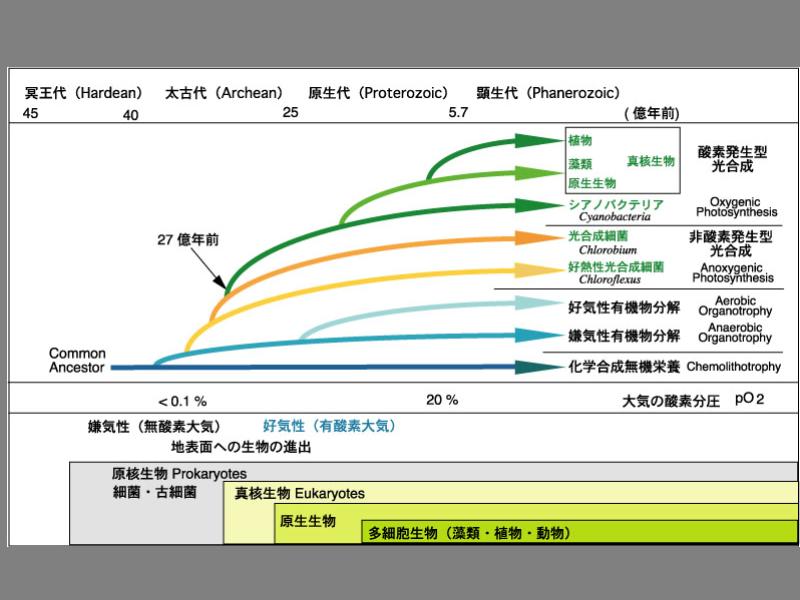
地球生命の進化時計
|

|
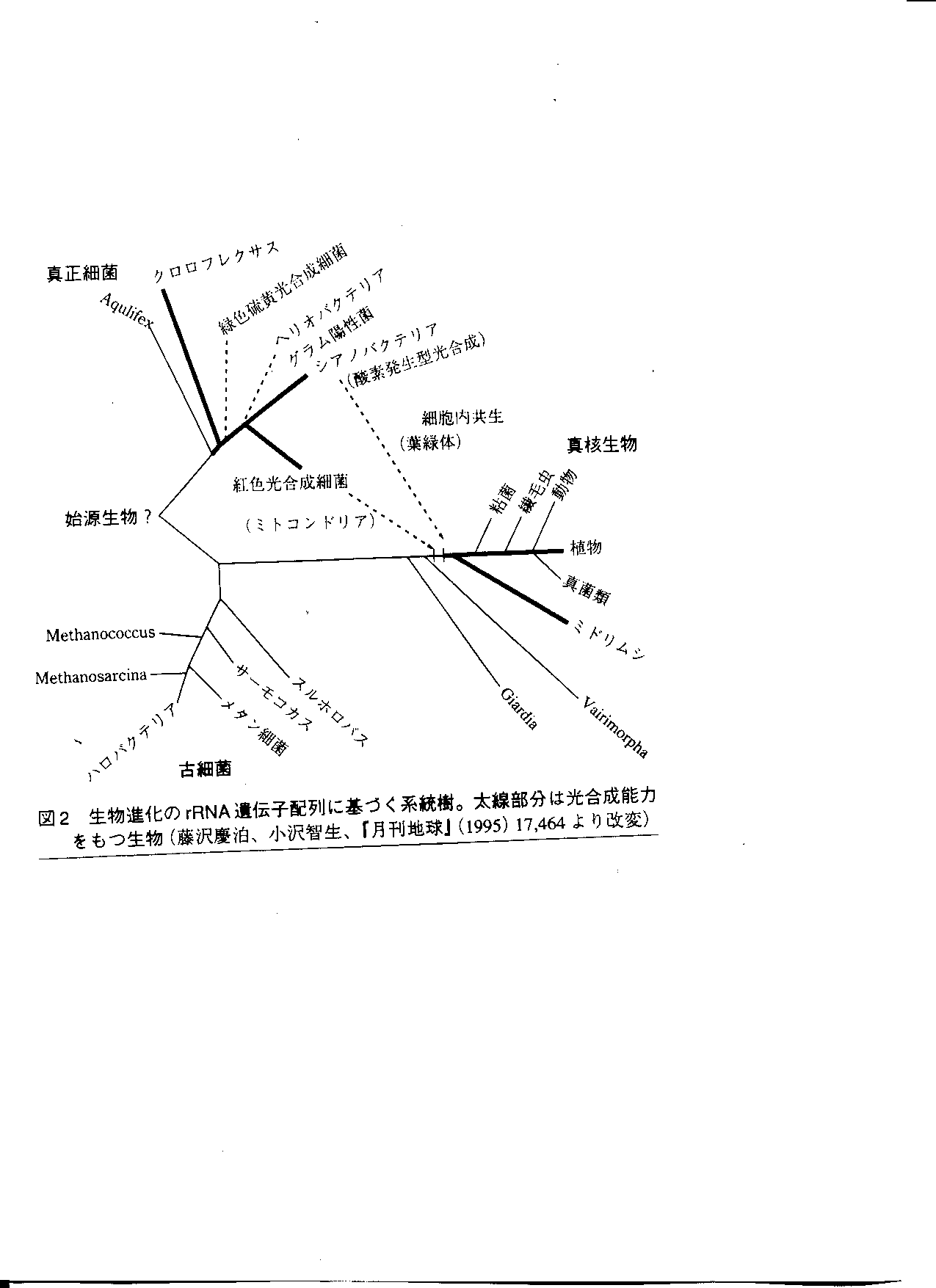
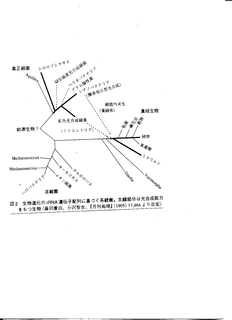
光合成に関する遺伝子による系統樹
|
|
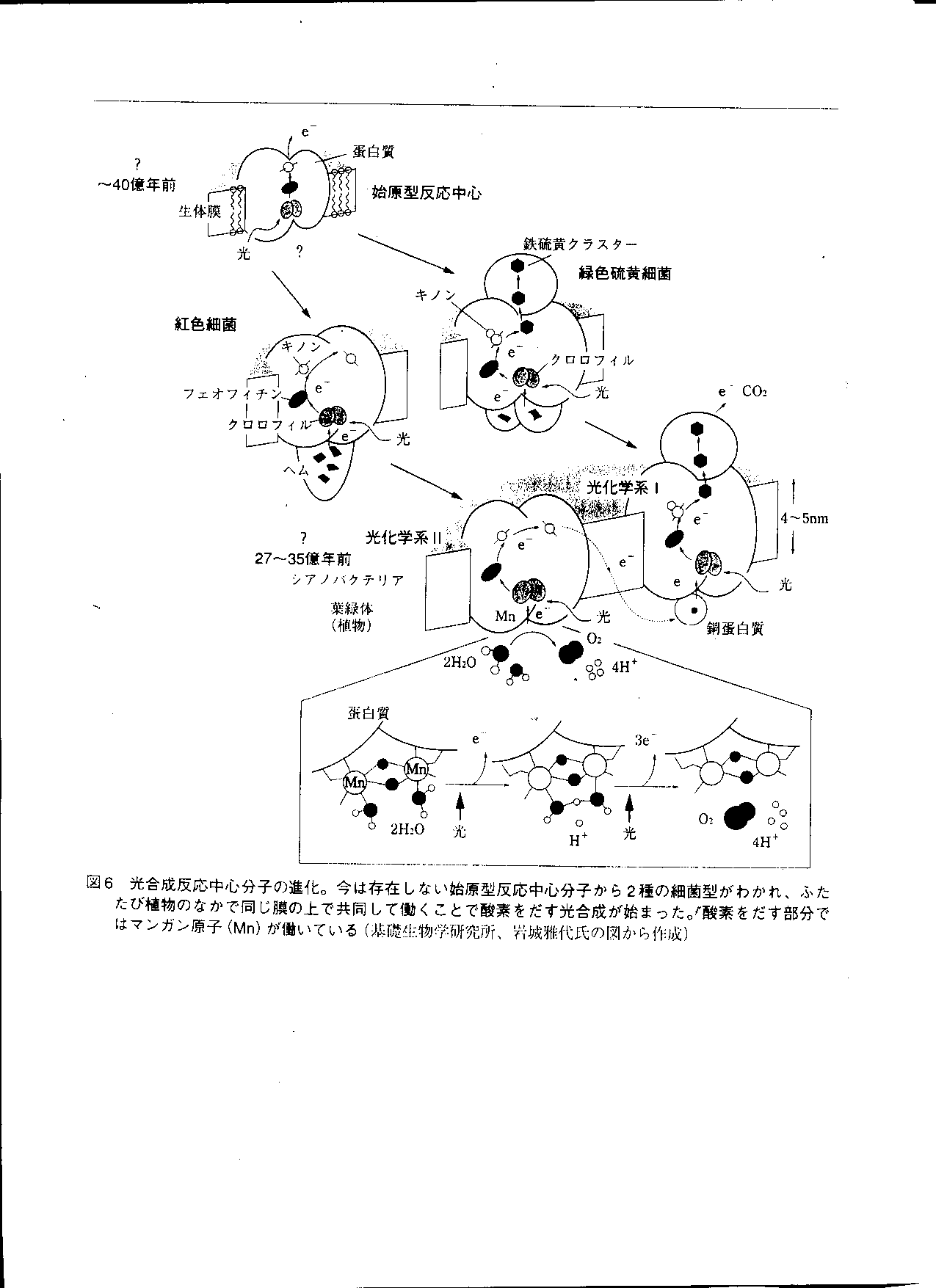
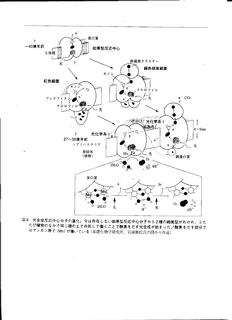
光合成反応中心の進化
- クロロフレクサス(緑色硫黄細菌)とクロロビュウム
(紅色細菌がそれぞれもっていた光合成反応中心が,
シアノバクテリア内で 1 つの反応中心に合体した.
- クロロフレクサスとクロロビュウムが持っていた反応中心では,
エネルギーの低い電子しか利用できない.
|
|
光合成細菌の系統樹上での位置
- シアノバクテリア内の反応中心が真核生物の葉緑体へ進化する.
|
|
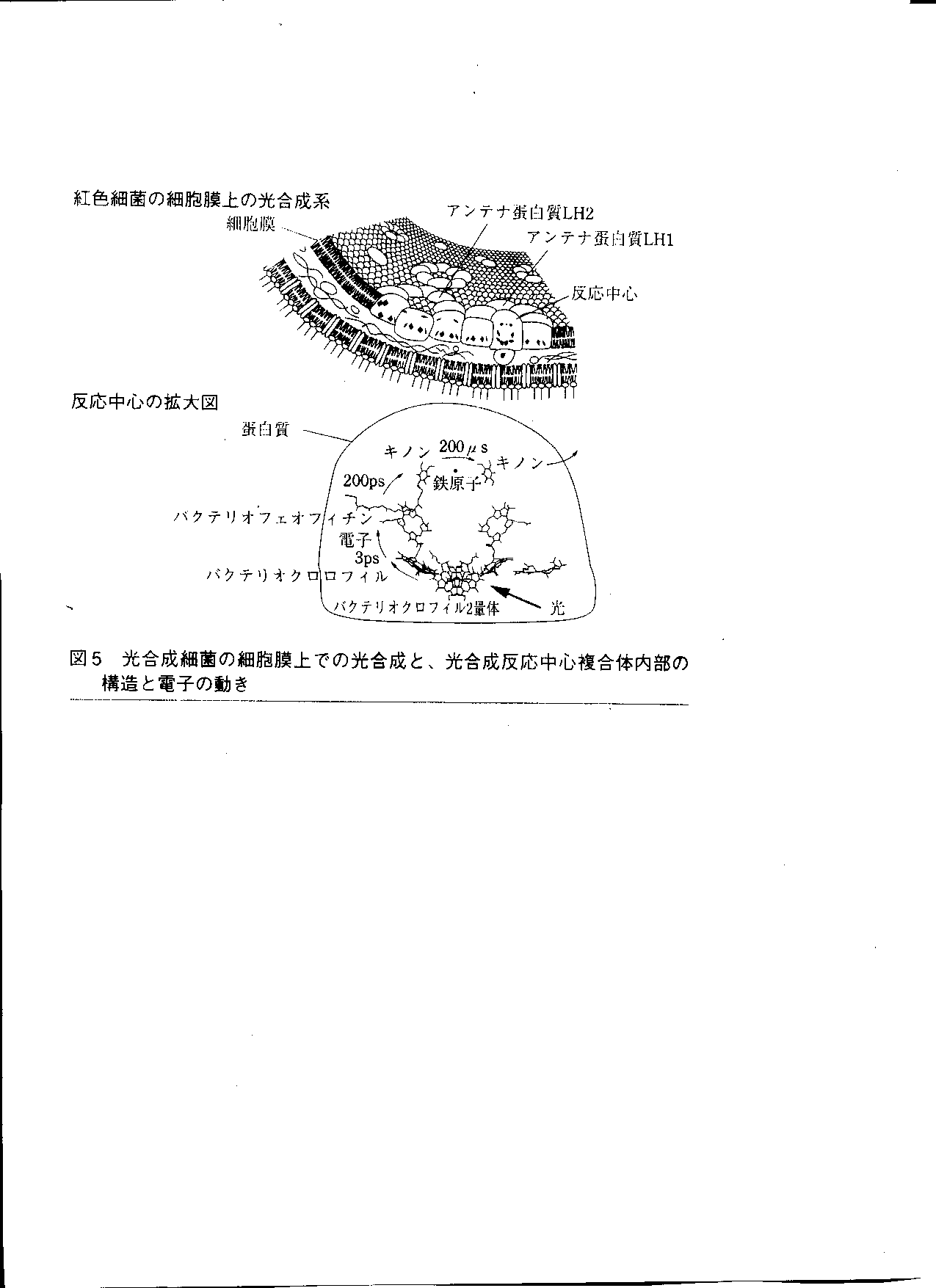
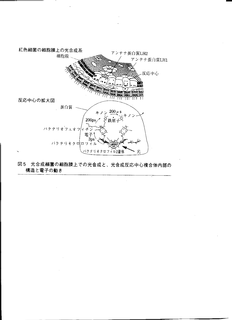
光合成反応中心の拡大図
- 反応中心の周囲に集光装置のようなアンテナタンパク質がある.
|

|
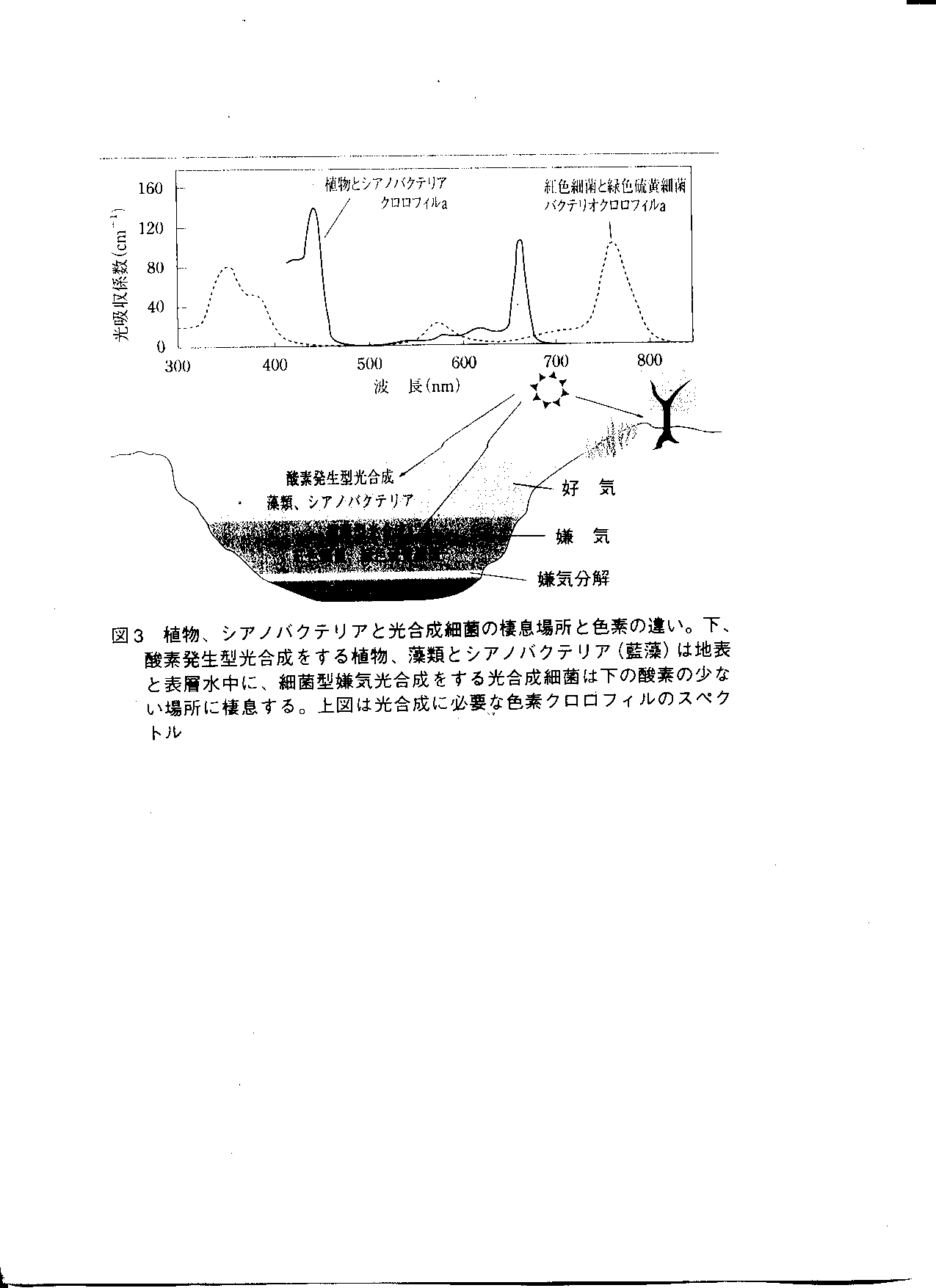
光合成時に用いる光の波長
|

|
ロドプシン: 光エネルギーを生体エネルギーに変換する物質
クロロフィル(葉緑体)とはことなる変換物質
|

|
系統樹: CO2 固定サイクル
- CO2 固定サイクルはカルビン-ベンソンサイクル(普通の植物の炭素固
定サイクル)だけではない
- 逆 TCA サイクルは -COOH 基に CO2 をつなげるサイクル
(動物の有機物分解の逆サイクル)
|

|
バクテリアの進化と地球史との関係
|

|
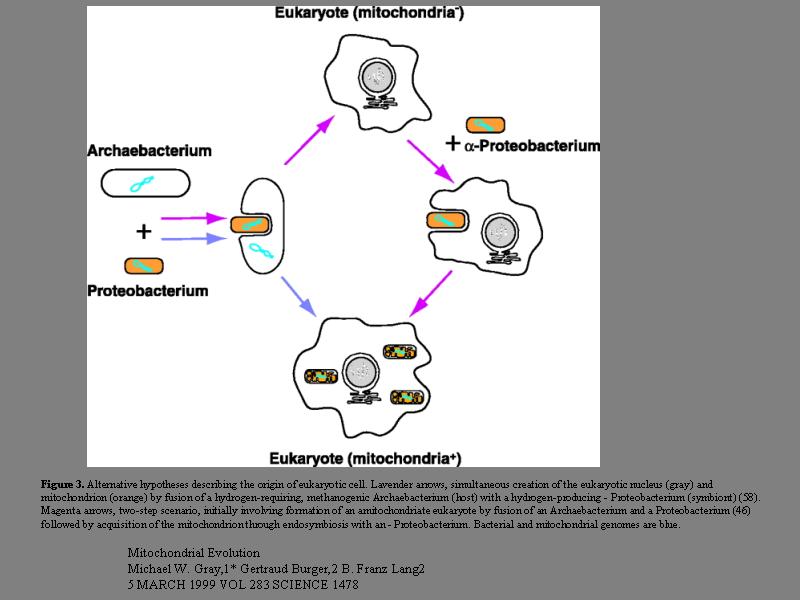
ミトコンドリアの取り込み
|

|
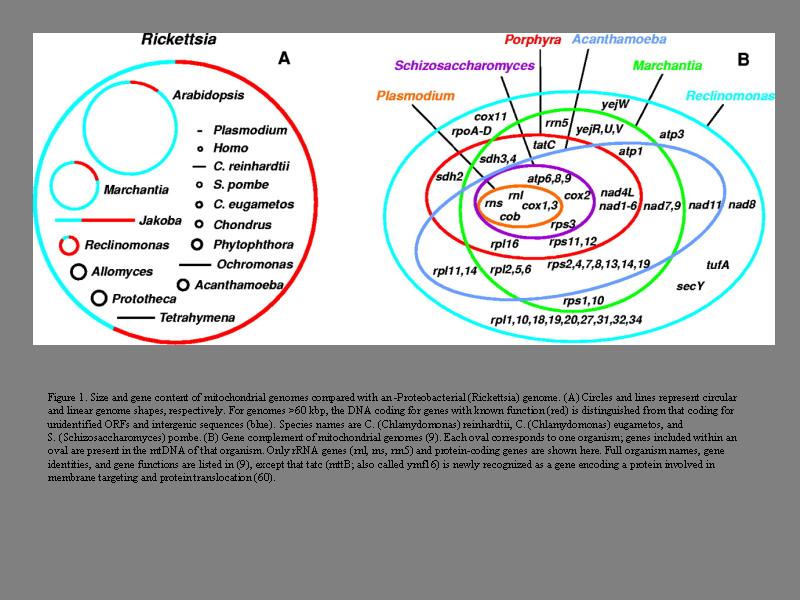
いろいろミトコンドリアの遺伝子の長さ
|
|
|
ミトコンドリアの系統樹上の場所
|

|
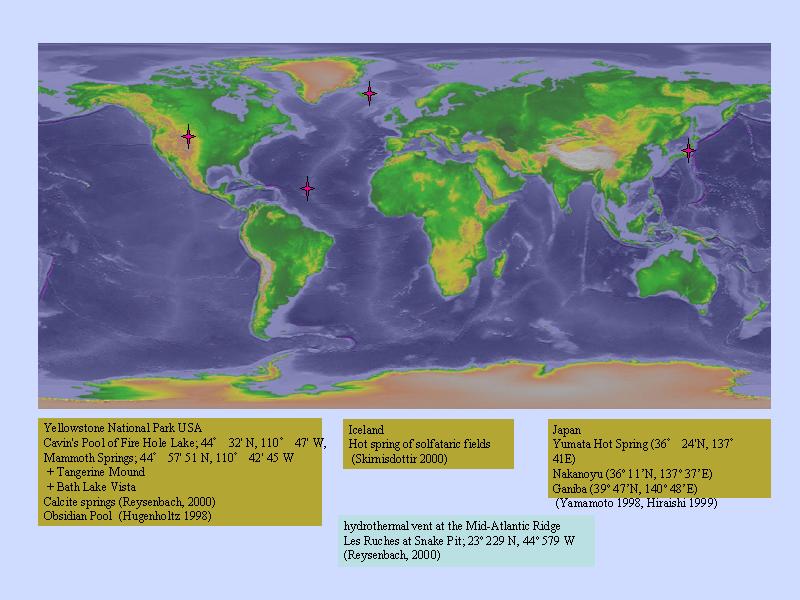
バイオマットの分布
- イエローストーン
- 日本
- アイスランド
- オーストラリア・ニュージーランド(未確認)
|

|
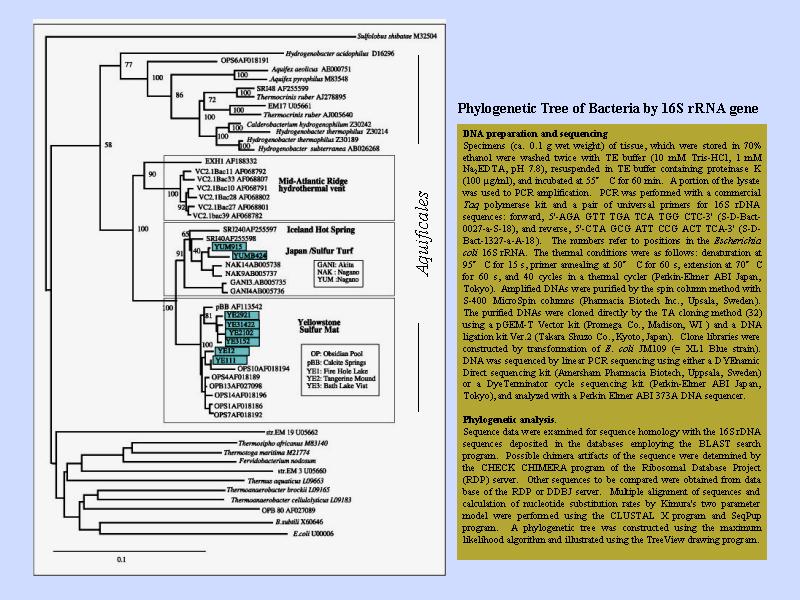
イオウシバの系統樹上の分布
環境に非常に影響されるため, 生息地域である程度分類できる.
- 大西洋中央海嶺
- イエローストーン
- アイスランドと日本
図中分岐点の数字は検定確率を表す.
横方向の枝の長さは「世代交替時間」の相対的長さ. 絶対時間ではない.
世代交替時間が異なる場合, 長さの比較は意味を持たない.
|

|
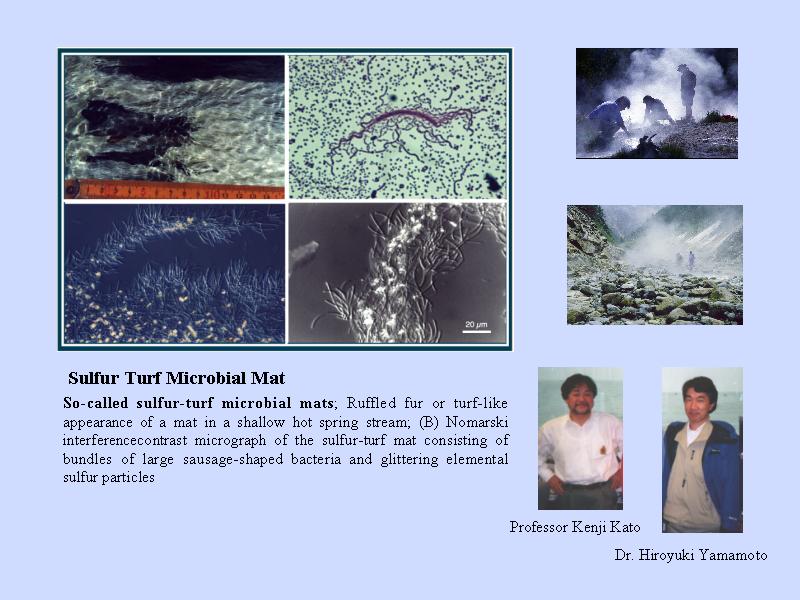
イオウシバの写真
|

|
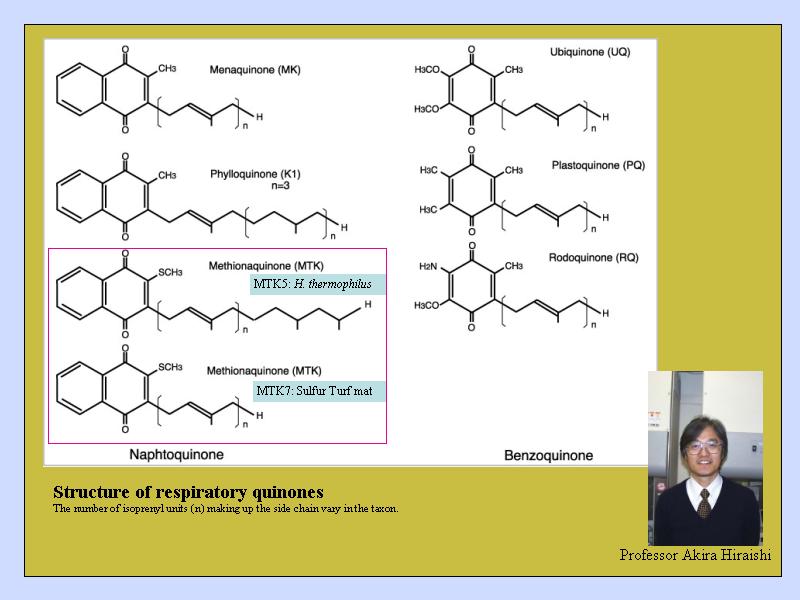
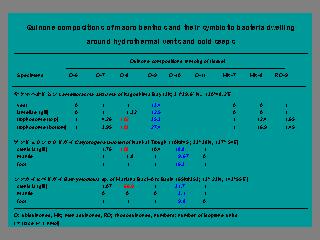
染色物質として用いるキノンの化学式
|

|
温泉バイオマット
なぜ温泉に注目するか?
- 海底の熱水噴出孔では, 多細胞生物がすでに進出している.
- 好熱性細菌は噴出孔のごく付近にしか存在しない.
- 温泉の方が原始的環境を観察しやすい.
- 温泉バイオマットのある場所には細菌しか存在しない.
温泉バイオマットは温度によって種類が異なる.
|

|
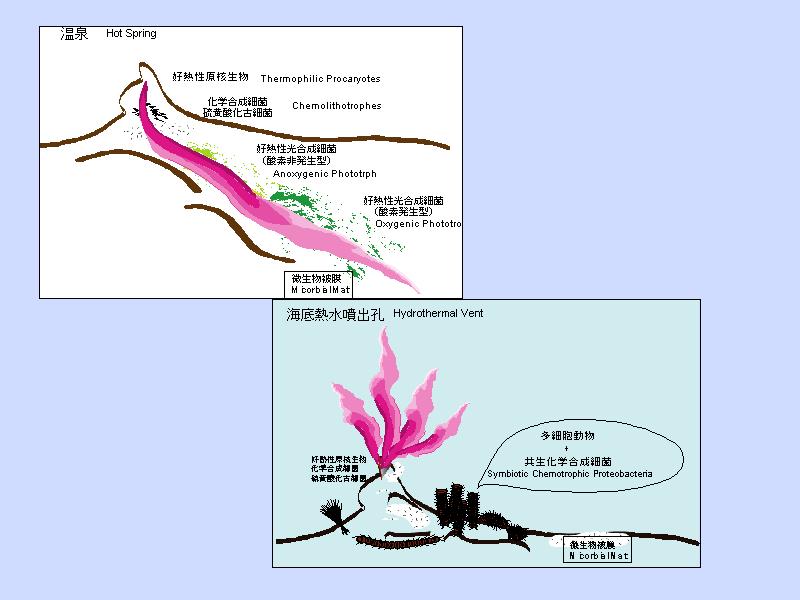
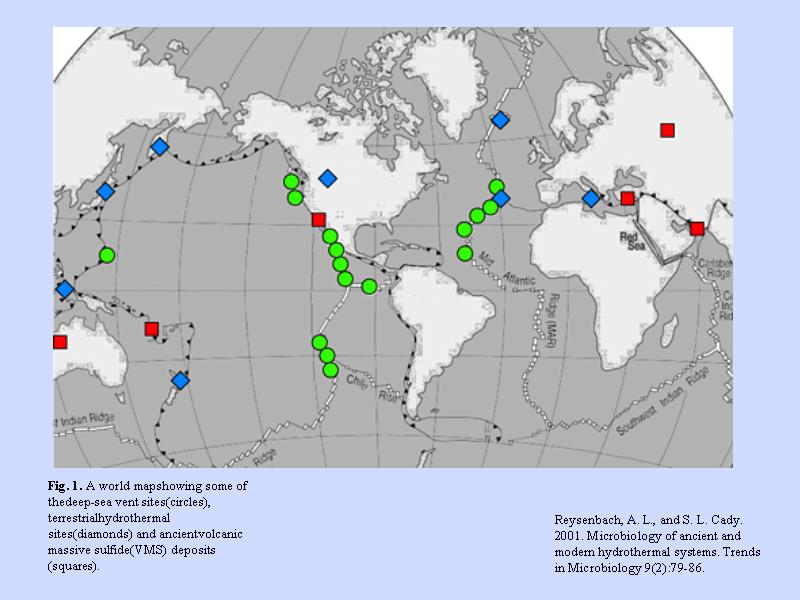
生物のいる温泉(熱水)分布
|

|
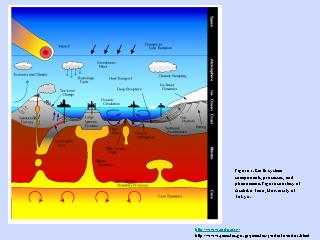
熱水系の模式図
|

|
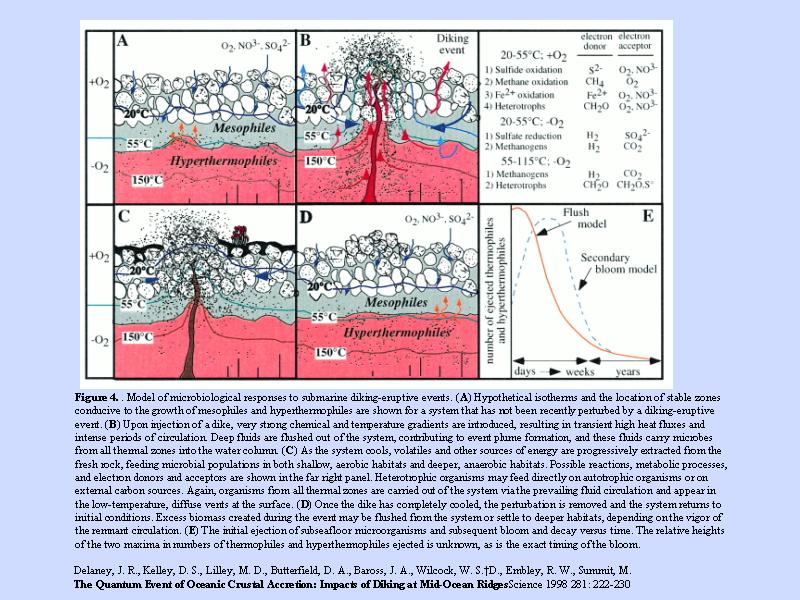
熱水系の断面 (1)
|

|
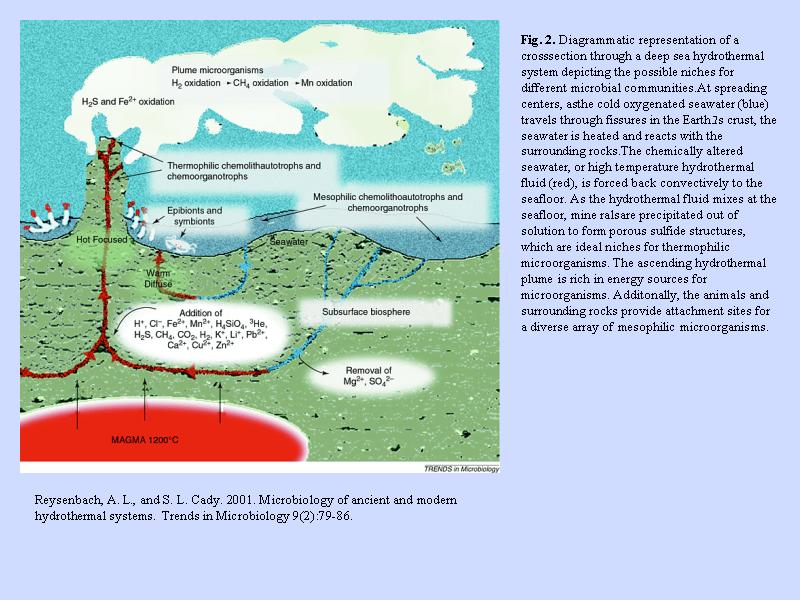
熱水系の断面 (2)
- 熱水は海水が循環することで供給される
- 熱水中の鉱物成分は循環路のある海底岩石の成分を反映
|

|
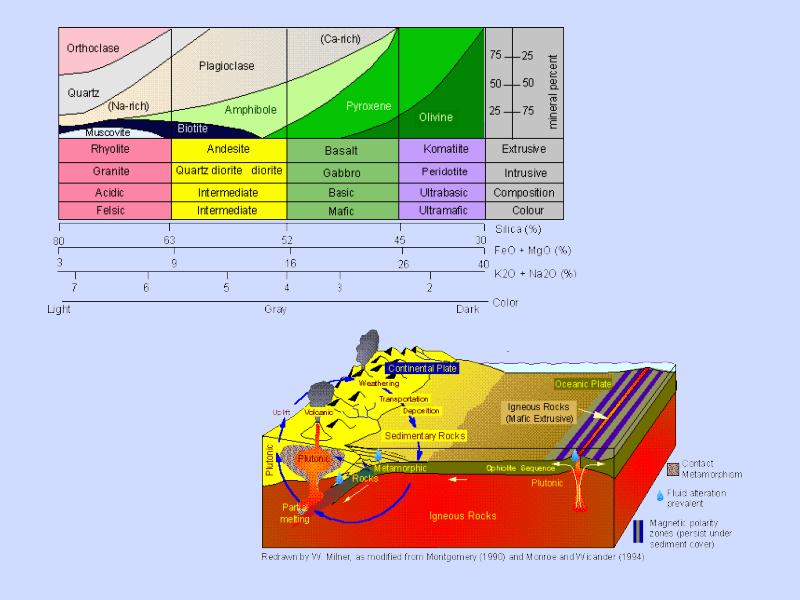
岩石の分類
|
|
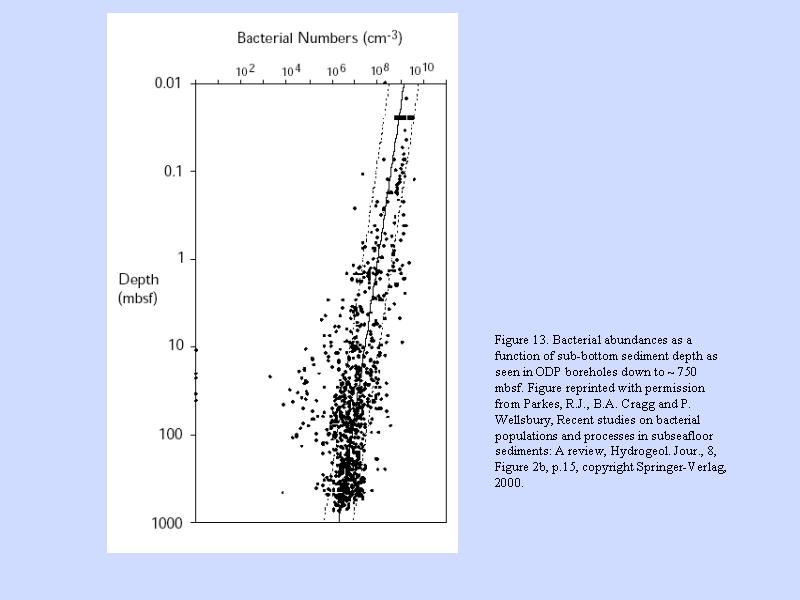
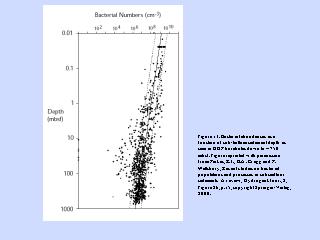
海底のバクテリア深度分布
|

|
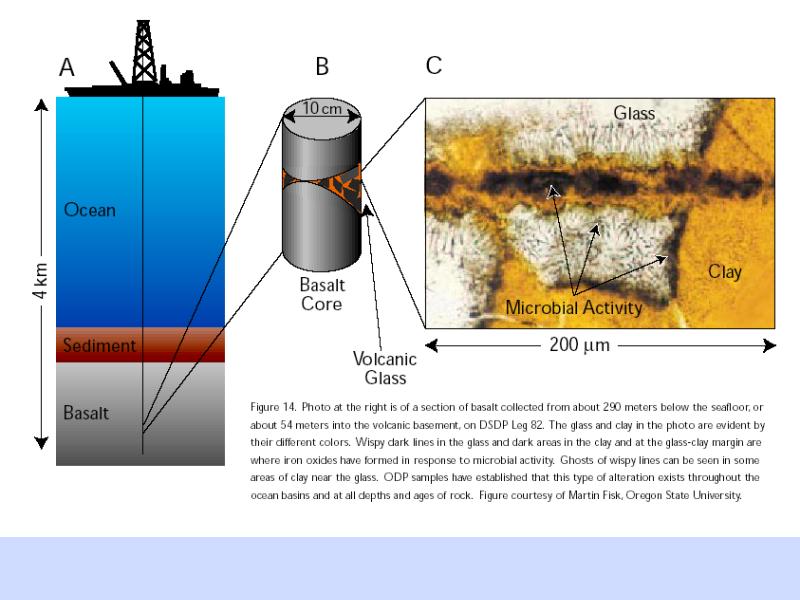
海底サンプリング
|
|
OD21 の模式図
|

|
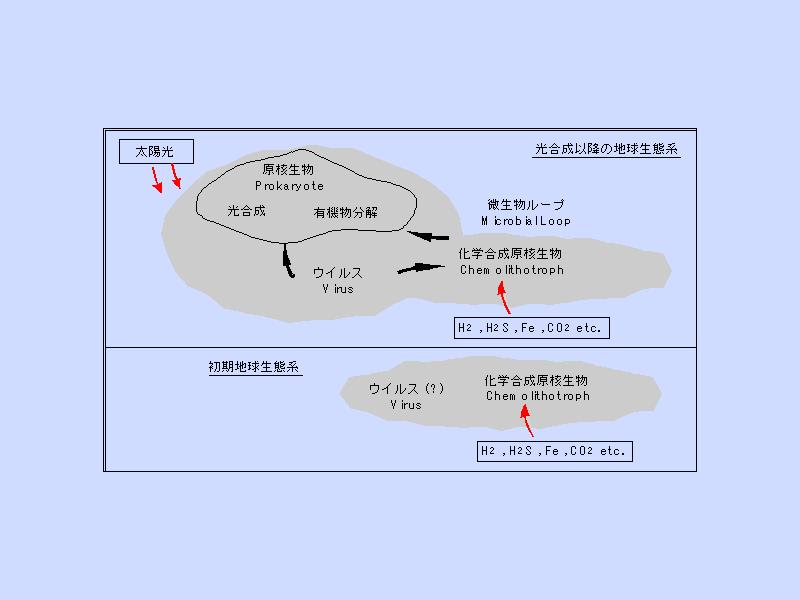
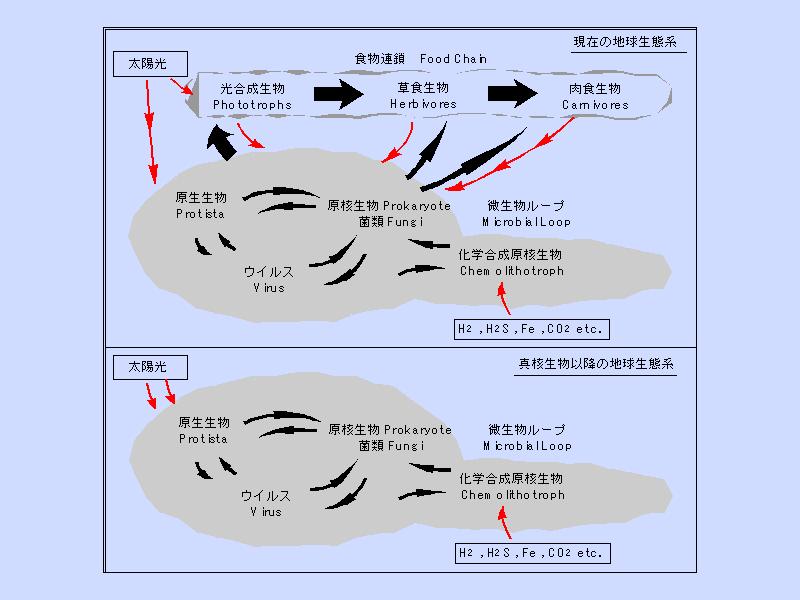
生態系の進化 (1)
- 化学合成生物だけが存在する時の生物生産量はあまり多くない
- 光合成を獲得することで生物生産量は急増する
|

|
生態系の進化 (2)
|
|
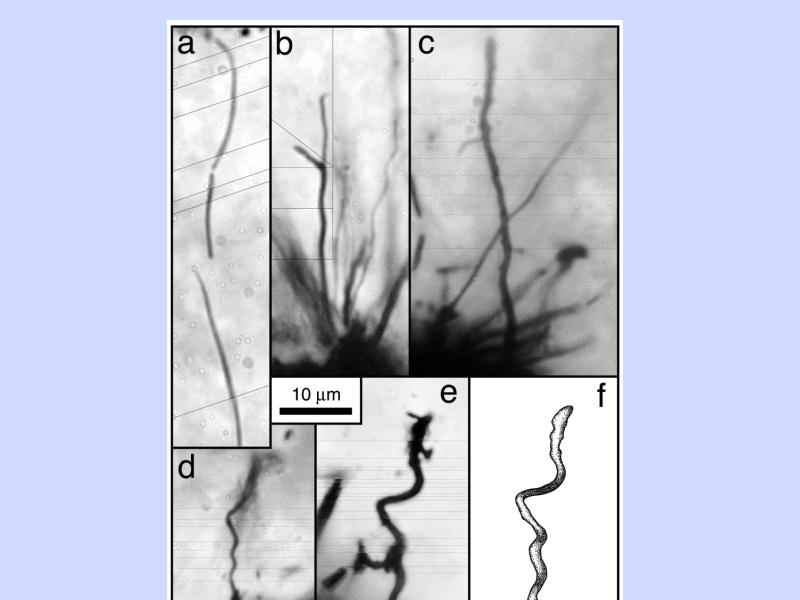
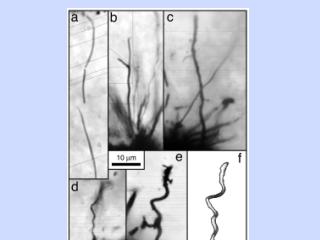
微生物の化石 (1)
|

|
微生物の化石 (2)
|

|
太古代の生態系の模式図
|

|
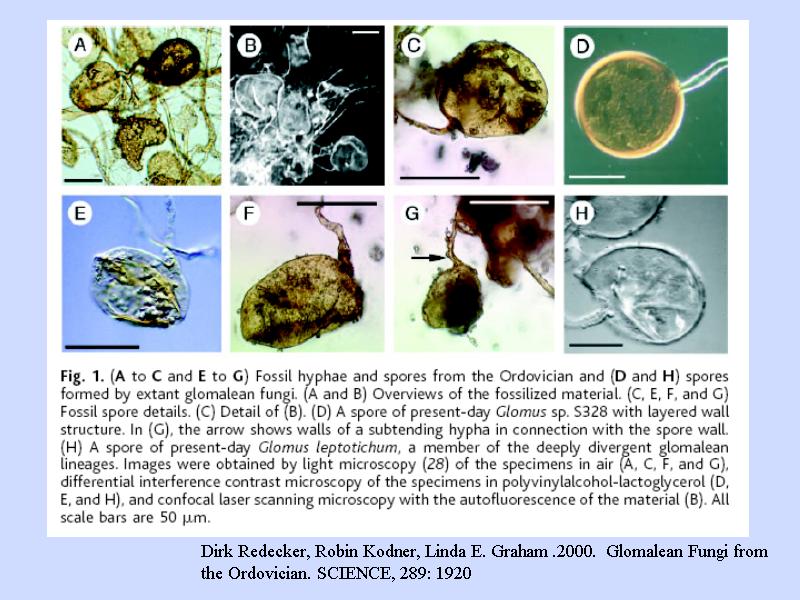
カビの化石
- 植物が上陸した時代の化石
- 植物の根に付着 (共生) していた
- 植物はカビを根に付けて, ともに上陸したと考えられている
|
Odaka Masatsugu
2001-09-11
|